Biology The Study of Life: Part 17 (Genes and Chromosomes)

Introduction
This informative series of posts will explore modern biology; the fundamental principles of how living systems work. This material will always be presented at the level of a first-year college biology course, without assuming any prior background in biology or science. It also presents material in a conceptual format. Emphasizing the importance of broad, unifying principles, facts and details in the context of developing an overarching framework. Finally, the series takes a historical approach wherever possible. Explaining how key experiments and observations led to our current state of knowledge and introducing many of the people responsible for creating the modern science of biology.
This post extends the discussion of how the movement of DNA explains patterns of trait expression.
First observed by Mendel in the middle of the 19th century and moves to a more modern understanding of the cellular and molecular basis of genetics that began at the turn of the 20th century. It covers Thomas Hunt Morgan’s discovery of the fact that some traits are more likely to be transmitted to offspring of one sex than the other. This discovery provided a key piece of evidence that chromosomes are the cellular structures responsible for trait transmission, and it played a key role in the reemergence of Mendel’s work, which had been virtually ignored for decades.
Mendel’s laws are consistent with what we now know about the movements of chromosomes during gamete formation.

Mendel himself knew nothing of this and his work was largely forgotten for a generation or more. Biologists in the second half of the 19th century began to observe dark bodies in the nuclei of cells before they divided. Walter Sutton and Theodor Boveri independently observed that the movement of these bodies reduced the number of bodies by half during gamete formation and realized that this might explain the patterns of trait inheritance Mendel had described. Sutton and Boveri published their ideas in 1903, introducing the Chromosomal Theory of Inheritance, in which each gene has its own location, or locus, on a chromosome. Genes can then independently assort, producing the patterns Mendel observed.
Thomas Hunt Morgan, working at the turn of the 20th century, was interested in mechanisms of inheritance and set out to find a suitable living system on which to work.

Morgan was especially interested in how mutations arose and wished to study this phenomenon in guinea pigs but could not obtain funding for this work. Morgan began to work with fruit flies (Drosophila melanogaster), which have since become the dominant model system for understanding genetics at all levels. Fruit flies have many advantages. They are cheap and easy to obtain. They are easy to raise in the laboratory. They have a very short generation time, allowing experimental crosses to be done in a short period of time. Fruit flies had a problem, however; they have no obvious phenotypic variations. To remedy this, Morgan attempted to create his own variants by exposing flies to agents known to induce mutations.
Morgan’s terminology has become the standard for many geneticists.

{kind=link}
Morgan and his colleagues referred to fruit flies found in nature as having wild-type phenotypes. Any flies with unusual phenotypic characteristics were referred to as mutant phenotypes, based on the assumption that the characteristic resulted from a mutation in an allele. Morgan and later fruit fly geneticists labeled alleles in a slightly different way than we have seen. For example, a reduction in wing size is called the vestigial wing mutation, and the mutant allele is labeled vg. The wild-type allele for that gene would then be vg+.
For any mutation we examine, we can assume that a wild-type fly is homozygous for the wild-type allele.
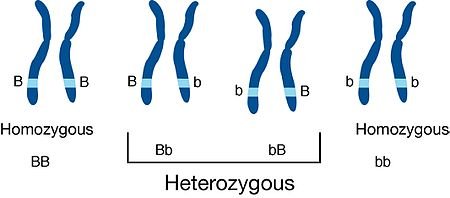
Mutants, therefore, are at least heterozygous for the mutant trait. Normally, the wild-type allele is dominant; thus, mutants often are homozygous for the recessive mutant allele. Most mutations make alleles dysfunctional. For example, in pea plants, the dominant purple-flower allele might code for a purple pigment protein, whereas the recessive white allele codes not for a white pigment but for no pigment at all.
After many failed attempts to induce mutations, an undergraduate working with Morgan was washing the milk bottles used to raise flies and noticed a single male fly in one bottle that was unlike all the rest.

Instead of red eyes, it had white eyes. This fly eventually enabled Morgan and his colleagues to prove that genes reside on chromosomes. When Morgan crossed this mutant white-eyed male with a wild-type red-eyed female, all the F1 offspring had red eyes, just as we would predict if the red-eye allele is dominant. When Morgan crossed F1 individuals, he found the expected 3:1 ratio of red eyes to white eyes in the F2 generation. However, all the female flies had red eyes, while half the males had red eyes and half had white eyes. That is, the allele for white eyes seemed to transmit only to male offspring. A sex-linked trait is one whose expression depends on the sex of the organism.
Morgan showed that females could have white eyes, but they needed two copies of the white-eye allele, while males needed only one copy. Morgan proposed that the white-eye gene must occur on the sex chromosomes, a unique pair of chromosomes that comprises a large X chromosome and a smaller Y chromosome that carries few functional alleles. If the eye-color gene is located on the X chromosome, females (which are XX in flies, as in humans) carry two alleles, while males (XY) carry just one. Males are referred to as hemizygous in this respect, because many genes on the X chromosome can occur only once in them.
Sex-linked traits explain F1 generation genotypes and phenotypes.
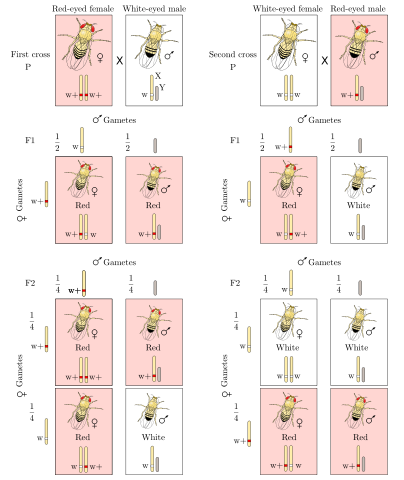
All F1 females have red eyes but are heterozygous because they receive one X chromosome from the mother and one from the father. Because the white-eyed father has only one X chromosome, it can transmit only the white-eye allele to all of its offspring. All F1 males carry the wild-type allele because they must receive a Y chromosome from the father and an X chromosome from the mother. As expected, sex -linked traits also predict F2 generation genotypes and phenotypes. Because F1 females are heterozygous, half their gametes will have the wild-type red allele and half will have the mutant white allele. Because F2 males are hemizygous and get their single allele from the mother, half of them will have red eyes and half will have white eyes. The X chromosome that F2 females receive from their fathers will always have the wild-type allele. Because this allele is dominant, F2 females will have red eyes, regardless of which allele is in the gamete from the mother. Morgan was then able to unambiguously connect the distribution of sex -linked phenotypic traits to allele distribution, proving that genes actually resided on sex chromosomes (and, therefore, on all other chromosomes).
END PART 17
BIOLOGY THE STUDY OF LIFE:
PART 1 INTRODUCTION
PART 2 WHAT IS LIFE
PART 3 ORIGIN OF LIFE
PART 4 CELL TO ORGANISM
PART 5 PROTEINS
PART 6 CODE OF LIFE
PART 7 DOUBLE HELIX
PART 8 REPLICATING DNA
PART 9 CENTRAL DOGMA
PART 10 GENETIC CODE
PART 11 DNA TO RNA
PART 12 RNA TO PROTEINS
PART 13 MISTAKES HAPPEN
PART 14 WHEN CELLS DIVIDE
PART 15 MENDEL'S PEA PLANTS
PART 16 SEX AND VARIATION
PART 17 GENES AND CHROMOSOMES

 or
or  @pjheinz
@pjheinzImage Credits:
ALL IMAGES UNLESS NOTED - Wikipedia
Thanx for this posts. Waiting for next :-)
Thanks for reading them, I hope you find the content informative and accessible.