Estudio de los factores intensidad lumínica, concentración de bicarbonato sódico y temperatura sobre la velocidad fotosintética en individuos (Phaseolus vulgaris) cultivados en sol y sombra.
Hoy les traigo una especie de guía (que intentaré convertir en compendio de actividades) con un par de "experiencias" sobre fisiología vegetal con enfoque principal a la docencia, mediante la práctica, para la enseñanza pedagógica y didáctica de las características primordiales del proceso llamado fotosíntesis y de los factores que lo afectan.
Las plantas utilizan la radiación solar para sintetizar compuestos orgánicos a partir del CO2 atmosférico, agua y nutrientes del suelo. La cantidad de carbono fijado en la fotosíntesis es controlada principalmente por la radiación y la temperatura y es limitada por la disponibilidad de agua y CO2. Como objetivo principal se estudiaron los efectos de factores externos sobre la velocidad fotosintética. Se prepararon las muestras obteniendo 20 discos de hojas de Phaseolus vulgaris, luego se colocaron en un tubo de ensayo con 10 mL de solución amortiguador citrato-fosfato o de bicarbonato de sodio y se infiltraron con una bomba de vacío. Posteriormente se colocaron 10 discos en una cápsula de Petri con 15 ml de la misma solución (duplicado) y se colocaron bajo una fuente de luz. Se midió el tiempo que tardó cada disco en subir y se determinó la velocidad fotosintética. Los parámetros variados fueron intensidad lumínica, disponibilidad de CO2 y temperatura, además se trabajó con plantas cultivadas en sol y en sombra. Se obtuvo máximos de velocidad a 5W/m2 en ambos tipos de plantas y mínimos a 15W/m2, en el caso de la planta de sombra muestra un pico de velocidad en la concentración 10 μM de NaHCO3 (0,004 cm/s), para luego disminuir y volverse más o menos constante, por su parte la planta de sol mostro un pico en 20 μM de NaHCO3 (0,007 cm/s), finalmente la de sombra muestra un aumento poco marcado de la V.F hasta los 30 °C, para luego alcanzar un mínimo de V.F a los 35°C.
INTRODUCCIÓN
La fotosíntesis es un proceso en virtud del cual los organismos con clorofila, como las plantas verdes, las algas y algunas bacterias, capturan energía en forma de luz y la transforman en energía química. La fotosíntesis se realiza en dos etapas: una serie de reacciones que dependen de la luz y son independientes de la temperatura, y otra serie que dependen de la temperatura y son independientes de la luz. La velocidad de la primera etapa, llamada reacción lumínica, aumenta con la intensidad luminosa (dentro de ciertos límites), pero no con la temperatura. En la segunda etapa, llamada reacción en la oscuridad, la velocidad aumenta con la temperatura (dentro de ciertos límites), pero no con la intensidad luminosa (Bernstein & Nieman, 1960).
Las plantas de sol tienen hojas pequeñas y numerosos estomas, el mesófilo fotosintético está integrado por muchas capas (lo que disminuye su relación superficie /volumen) y utilizan el nitrógeno para la regeneración de la RuBisCO, además tienen más cantidad de xantofilas. Las xantofilas se utilizan para disipar el exceso de energía y aunque generalmente en las plantas sus niveles son relativamente constantes, en las plantas perennes se ven importantes fluctuaciones en su concentración siendo llamativamente elevada durante el invierno. Otra excepción a esto la constituyen las plantas que viven en ambientes sombríos pero que pueden recibir mucha radiación en muy poco tiempo. Las plantas de sombra, tienen por lo general las hojas más grandes y menos estomas y el mesófilo fotosintético no está tan desarrollado como en las de sol, por lo que su relación superficie /volumen es alta. Sus cloroplastos tienen más membranas apiladas por lo que tienen más PSII que PSI, esto es debido a que a las plantas de sombra les llega más luz de 700nm y el PSII funciona a 700nm. Los estudios indican que aproximadamente hay 3 PSII por cada PSI. En las plantas de sombra, el nitrógeno no es empleado para la regeneración de la RuBisCO sino para la síntesis de proteínas relacionas con el complejo antena, donde hay más clorofila B (Awashti, 2005; Erickson, 1992).
La luz con intensidad elevada daña el proceso de la fotosíntesis. La fotoinhibición tiene lugar cuando la intensidad de la luz excede la capacidad del ciclo del carbono en la planta. La saturación de la iluminación destruye los pigmentos de las plantas, impidiendo la conversión suficiente de energía. Las plantas tienen mecanismos para combatir los daños asociados con la elevada intensidad de la luz, como la disipación de la energía, la desactivación fluorescente y pigmentos especializados de clorofila (Meléndez et al., 2001).
Por lo que los objetivos de la investigación fueron:
• Estimar la velocidad fotosintética en discos de hoja en relación a la intensidad de luz.
• Estimar la velocidad fotosintética en discos de hoja en relación a la temperatura.
• Estimar la velocidad fotosintética en discos de hoja en relación a la disponibilidad de CO2.
MATERIALES Y MÉTODOS
Para ambas experiencias se procedió a obtener discos de hojas de Phaseolus vulgaris mediante un sacabocados. Al obtener 20 discos de hojas se colocaron en un tubo de ensayo con 10 mL de amortiguador citrato-fosfato (en evaluación del efecto de la intensidad de luz en la velocidad de fotosíntesis) o de bicarbonato de sodio (en evaluación del efecto de la temperatura en la velocidad de fotosíntesis) y se infiltraron con una bomba de vacío, agitando el tubo y destapando el mismo. Posteriormente se desechó el líquido de infiltración y se colocaron 10 discos en una cápsula de Petri colocándolos con una distribución homogénea con 15 ml de la misma solución, luego se colocó en otra capsula el duplicado y se procedió a colocar bajo una fuente de luz. Finalmente se midió el tiempo (en segundos) que tardó cada disco foliar en subir a la superficie. Se tomaron los tiempos incluidos los duplicados, se promediaron y se determinó la velocidad de fotosíntesis. Las diferencias principales entre las actividades realizadas fueron los parámetros que fueron variados, en la experiencia 1 se modificó la intensidad lumínica (0; 5; 10; 15; 20 y 25 W/m2), en la 2 se modificó la temperatura (15; 25; 30; 35; 40 y 45 °C) y con una intensidad lumínica constante de 25W/m2, y en la 3 se evaluó el efecto de la disponibilidad de CO2 en la velocidad de fotosíntesis (0; 5; 10; 20; 50 y 100 μM), además en cada caso se trabajó con plantas cultivadas en sol y en sombra.
RESULTADOS Y DISCUSIÓN.
EXPERIENCIA #1. Evaluación del efecto de la intensidad de luz en la velocidad de fotosíntesis.
En líneas generales se observó un comportamiento similar entre plantas de luz y de sombra, con un valor máximo de velocidad promedio en plantas de sombra a 25W/m2 (aproximadamente 0,0045cm/s), se puede evidenciar más específicamente la distribución en plantas de luz y sombra en las siguientes figuras 1 y 2.
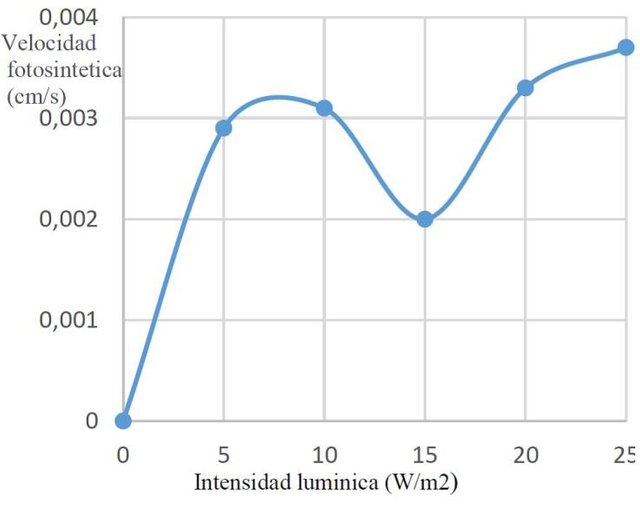
Figura 1. Velocidad fotosintética en plantas de sol a diferentes flujos protónicos.
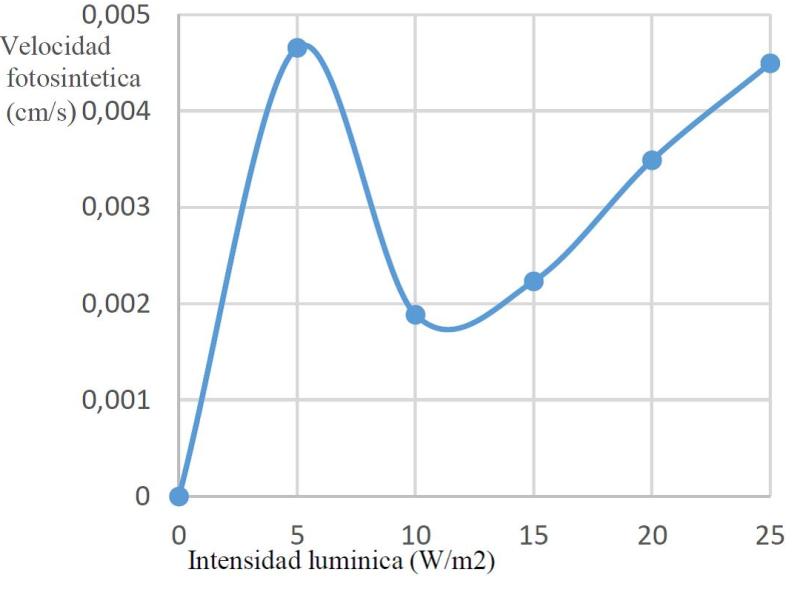
Figura 2. Velocidad fotosintética en plantas de sombra a diferentes flujos protónicos.
En oscuridad no hay fotosíntesis, lo cual no quiere decir que no se respire; es por esto que se necesita un valor mínimo de irradiancia para el cual la tasa de consumo de CO2 y la de desprendimiento por respiración sean iguales. En ese punto la tasa de asimilación de CO2 fotosintético es 0 y se le llama punto de compensación de luz. Cuanto más aumente la irradiancia más aumenta la fotosíntesis hasta alcanzar el punto de saturación consecuencia de la limitación impuesta por la velocidad de las reacciones del carbono, a partir de ese punto la fotosíntesis no aumenta con la intensidad lumínica. Es el punto de saturación de luz. Las plantas de sombra se saturan a irradiancias más bajas que las de sol y de igual modo, su punto de compensación es también menor, esto es debido a que su tasa de respiración también es menor. Al comparar pantas de sombra y pantas de sol se observa que las de sombra se saturan a irradiancias de 1000 μmol y las de sol a valores mucho más altos. La explicación a esto se encuentra en los requerimientos energéticos que tienen las plantas de sol, los cuales son mayores (Awashti, 2005; Herrera, 2011).
La luz es un factor limitante y a la vez dañino, por esto el aparato fotosintético cuenta con toda una serie de mecanismo de protección o defensa: mecanismos de supresión (disipación por calor), mecanismos de detoxificación de compuesto reactivos y por último, mecanismos de reparación del fotosistema. El ciclo de las xantofilas cobra especial importancia en este apartado. Consiste en la conversión de tres xantofilas que tienen distintos grados de epoxidación para producir la atenuación fluorescente de la clorofila por procesos distintos al fotoquímico (atenuación no fotoquímica). En presencia de luz intensa se favorece la conversión de violaxantina a zeaxantina. La zeaxantina se une a las proteínas de los complejos antena provocando cambios conformacionales que conducen a su desactivación y a la disipación de energía en forma de calor. La zeaxantina es más eficiente que la violaxantina. Si las reacciones lumínicas van más rápido que las del carbono se acidifica el lumen del tilacoides provocando la activando de la violaxantina de-epoxidasa; sintetizando zeaxantina y eliminando la excitación de la clorofila en forma de calor. Este mecanismo es común a todas las plantas. Es lógico pensar que las plantas de sol tengan más zeaxantina que las de sombra, ya que con más frecuencia sufrirán un exceso de luz (Meléndez & Molina, 2002).
EXPERIENCIA #2. Evaluación del efecto de la temperatura en la velocidad de fotosíntesis.
Como puede observarse en la figura 3, se obtuvo una velocidad fotosintética con varios picos en ambas condiciones lumínicas, la de sombra (figura 1. arriba) muestra un aumento poco marcado de la V.F hasta los 30 °C, para luego alcanzar un mínimo de V.F a los 35°C y luego continuar subiendo la velocidad de fotosíntesis, mostrando una relación casi directa (lo que se esperaba), por su parte las plantas de sol (figura 1. abajo) tiene un aumento brusco en la V.F entre los 25 y 30 °C disminuyendo y aumentando desde los 30 hasta los 40°C, respectivamente.
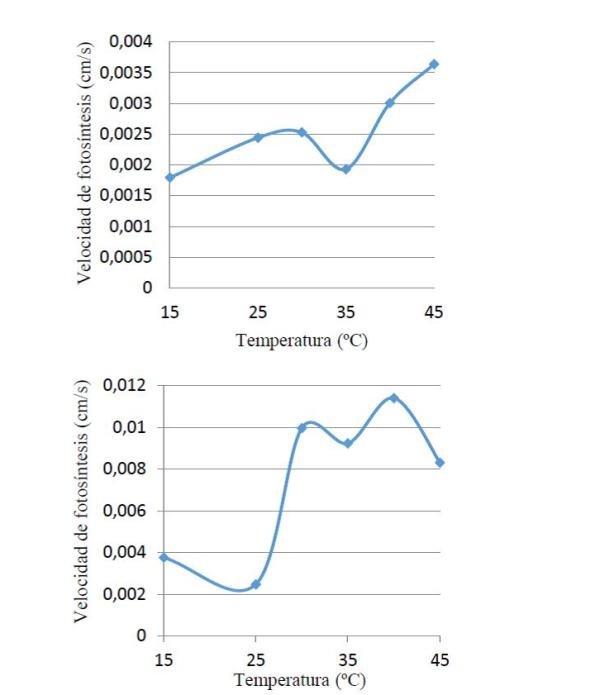
Figura 3. Velocidad fotosintética en plantas de sombra (arriba) y sol (abajo) a diferentes temperaturas.
La fotosíntesis es el proceso fisiológico más afectado por la temperatura alta; el daño causado puede llegar a depender de la estabilidad de los cloroplastos para mantener las reacciones fotosintéticas activas y de la velocidad de síntesis de la clorofila; así mismo, puede llegar a variar de acuerdo a la capacidad de respuesta del genotipo (Barrett & Gifford, 2005). Las hojas tienen una temperatura más alta que el ambiente por lo que cuentan con mecanismos de refrigeración tales como la emisión de luz a longitud de onda larga, fenómenos de conducción, convección y transpiración mediante la evaporación de agua en la superficie de la hoja.
Kimball (1978) señala que el efecto de las temperaturas altas causa, por un lado, un descontrol en los iones del agua, impidiendo el movimiento de solutos a través de las membranas de las células, y por otro, induce cambios en los procesos fisiológicos y bioquímicos incluyendo la fotosíntesis, respiración, actividad enzimática, estabilidad de la membrana celular y por último el crecimiento. Se ha observado en algunas plantas de cultivo que la fotosíntesis neta es inhibida a temperaturas arriba de 38ºC, aunque se ve más afectada cuando la temperatura se incrementa rápidamente. La principal causa de esto es la inactivación de la RuBisCO, la cual empieza a decrecer su estado de activación a temperaturas arriba de 32.5ºC y casi se inactiva completamente a 45ºC (Jungk, 1990). Otros estudios fisiológicos han demostrado que cuando aumenta la temperatura se produce una aceleración en la senescencia de las hojas, disminuyendo el período de actividad fotosintética (Miglietta & Raschi, 2003). Al respecto Drake et al. (1997) señalan que la tolerancia al calor está asociada a la característica de “stay-green”, la cual contribuye a mantener una mayor duración del aparato fotosintético.
La emisión de isopreno (compuesto de 5 carbonos y 2 dobles enlaces) que puede observarse en forma de neblina de color azul supone una pérdida de carbono que permite a la planta tolerar episodios controlados de aumento de temperatura estabilizando la membrana fotosintética y permitiendo la recuperación de la actividad fotosintética. Su emisión aumenta con la temperatura y con la luz y además favorece la formación de compuestos de efecto invernadero como el ozono. Es un mecanismo característico de plantas de sombra. Hay una serie de cambios en la velocidad fotosintética y en la eficiencia fotosintética en función de la temperatura. A partir de una determinada temperatura se favorece la actividad oxigenasa en detrimento de la carboxilasa. Esto se equilibra en mayor o menor medida aumentando la concentración de CO2 en torno a la RuBisCO. En plantas de sol el óptimo fisiológico se encuentra un valor de temperatura más alto que en las de sombra y además no se observa el aspecto negativo de la oxigenación con el aumento de la temperatura. En las plantas de sombra la eficiencia de la fotosíntesis disminuye al aumentar la temperatura mientras que en las plantas de sol no varía. Esa pérdida de eficiencia está relacionada con la fotorrespiración. Esto demuestra que las plantas de sol sólo son eficientes en ambientes cálidos y justifica el porcentaje tan reducido que representan. Cerca del ecuador son más eficientes las de sol, fuera de él lo son las de sombra (Fernández et al., 1999).
Otro factor importante para la fotosíntesis es la disponibilidad interna y externa de CO2 ya que el dióxido de carbono se combina con agua. El CO2 se transforma en azucares y otras moléculas altamente energéticas. En la figura 4 pueden observarse el comportamiento de velocidad fotosintética durante el aumento de la concentración del ion carbonato en la solución. En el caso de la planta de sombra muestra un pico de velocidad en la concentración 10 μM de NaHCO3 (0,004 cm/s), para luego disminuir y volverse más o menos constante, por su parte la planta de sol mostro un pico en 20 μM de NaHCO3 (0,007 cm/s) mostrando una relación indirecta entre V.F y concentración de NaHCO3 al aumentar la concentración a partir de 20 μM, como puede observarse en la figura 4.
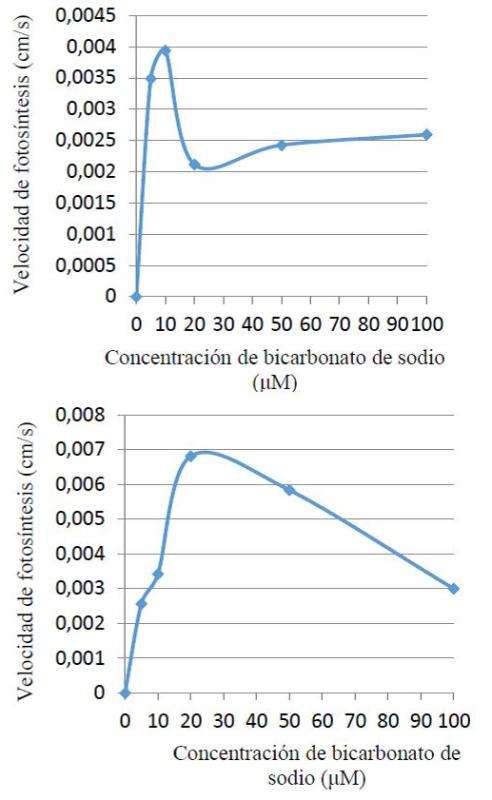
Figura 4. Velocidad fotosintética en plantas de sombra (arriba) y sol (abajo) a diferentes concentraciones de bicarbonato de sodio.
Se puede explicar el comportamiento de la fotosíntesis con respecto a la disponibilidad de CO2 mediante tres cinéticas. La primera, a bajas concentraciones de CO2, tiene tasas de fotosíntesis negativa porque se desprende CO2 a través de la respiración mitocondrial. Esta fase termina en el punto de compensación por CO2 (concentración de CO2 a la cual la fotosíntesis neta es igual a 0). Concentraciones de CO2 por encima del punto de compensación se inicia la segunda fase, es una cinética lineal que a medida que aumenta la cantidad de CO2 aumenta la fotosíntesis. La fase tres es la de saturación, llega un momento en que por más que aumente la concentración de CO2 la tasa de fotosíntesis no aumenta más, esto se debe a la capacidad de la planta de regeneración de la ribulosa-1, 5-bisfosfato (aceptor primario de CO2) (Fernández et al., 1998).
El efecto de la concentración de CO2 sobre la tasa de fotosíntesis neta en plantas de sombra y sol se da de la siguiente manera: La PEP carboxilasa de las plantas de sol tiene mucha más afinidad por el CO2 que la RuBisCO. Al tener un mecanismo de concentración de CO2, las plantas de sol en el momento en el que sube la concentración de CO2 atmosférica rápidamente se saturan. Los puntos de compensación de CO2 entre una planta de sombra y sol son distintos. La de sombra tiene más fotorrespiración de manera que para que la fotosíntesis neta sea igual a 0 tiene que aumentar más la concentración de CO2 atmosférico. La regulación descendente de la fotosíntesis debida a la [CO2] elevada consiste en un descenso de la tasa fotosintética desde valores iniciales que son superiores a los medidos en plantas a [CO2] ambiental hasta valores a [CO2] ambiental; se observa frecuentemente en experimentos en potes y ha sido atribuida a una limitación impuesta por el tamaño del sumidero, la disponibilidad de nutrientes y la restricción al crecimiento radical (Lawlor & Keys, 1993).
CONCLUSIONES
• La velocidad fotosintética en plantas de sol y sombra bajo condiciones de irradiación es máxima alrededor de los 5W/m2, mostrando esta última una pendiente mayor, es decir, estas se saturan más rápidamente.
• Los resultados obtenidos en la experiencia de la temperatura no concuerdan con los de Jungk (1990) ya que a temperaturas mayores de 32.5ºC aumenta gradualmente la velocidad de fotosíntesis tanto en plantas de sol como de sombra.
• La disponibilidad de CO2 mostró con respecto a la velocidad fotosintética dos de las tres fases cinéticas (“tasa fotosintética negativas” y la compensación) tanto en plantas de sol como en las de sombra, mas no se muestra un punto de saturación, se recomienda aumentar el número de muestras y la concentración de NaHCO3 para futuros estudios.
• Los errores en la medición de los parámetros estudiados y por ende las discrepancias con la bibliografía consultada pueden estar relacionadas a errores técnicos por parte de los investigadores.
REFERENCIAS BIBLIOGRÁFICAS
• Awashti, M. 2005. Rate of photosynthesis: limiting factors. Internacional Journal of Environmental Sciences. 2(3):201-206.
• Barrett, D. & Gifford, R. 2005. Acclimation of photosynthesis and growth by cotton to elevated temperature: interactions with severe phosphate deficiency and restricted rooting volume. Aust. J. Plant Physiol. 22:955-963.
• Bernstein, L. & Nieman, R. 1960. Photosynthesis, respiration and transpiration in green plants. Plant Physiology. 35: 589-598.
• Drake, B., González, M. & Long, S. 1997. More efficient plants: a consequence of rising atmospheric CO2. Annual Rev. Plant Physiol. Plant Mol. Biol. 48: 609-639.
• Erickson J. 1992. The greenhouse effect: tomorrow´s disaster today. McGraw-Hill, Inc., New York.
• Fernández, M., Pieters, A., Donoso, C., Tezara, W., Azkue, M., Herrera, C., Rengifo, E. & Herrera, A. 1998. Effects of a natural source of very high CO2 concentration on the leaf gas exchange, xylem water potential and stomatal characteristics of plants of Spatiphylum cannifolium and Bauhinia multinervia. New Phytol. 138: 689-697.
• Fernández, M., Pieters, P., Azkue, M., Rengifo, E., Tezara, W., Woodward F. & Herrera, A. 1999. Photosynthesis in plants of four tropical species growing under elevated CO2. Photosynthetica 37: 587-599.
• Herrera, L. 2011. The impact of light quality and leaf wetness on photosynthesis in north-west Andean tropical montane cloud forest. Plant Physiology Journal. 10: 111-117.
• Jungk, A., Asher, C. & Edwards, D. 1990. Influence of temperature on CO2 uptake kinectics of maize (Zea mays) and soybean (Glucine max). Plant Physiology. 14(2):233-252.
• Kimball, B. 1978. Temperature and agricultural yield: an assemblage and analysis of 430 prior observations. Agron. J. 75: 779-789.
• Meléndez, G. & Molina, E. 2002. El ciclo de las xantofilas, un indicador del estado fisiológico de los árboles. Ediciones del Laboratorio de suelos. Universidad de Costa Rica.
• Meléndez, L., Lizaso, J. & Ramírez, R. 2001. Factores reguladores del centro de reacción fotosintético y la tasa fotosintética en plantas verdes. Bioagro Rev. 13(3):111-116.
• Miglietta, F. & Raschi, A. 2003. Studying the effect of elevated CO2 and temperature in the open in a naturally enriched environment in Central Italy. Vegetatio 104(5): 391-393.
• Lawlor, D. & Keys, A. 1993. Understanding photosynthetic adaptation to changing climate. Journal of Plant Physiology. 29: 995-1003.
Interesante artículo, abordas un tema tan importante como lo es la fotosíntesis la cual es necesaria para la vida de los organismos de este planeta. Saludos.
Gracias, además de que sirve como protocolo para hacer experiencias propias para demostraciones a nivel de docencia por ejemplo, usando una especie vegetal ampliamente distribuida en Venezuela. Abrazo.